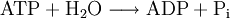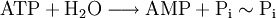Como la definición de Luz se da sólo a la radiación que causa sensación visual en el hombre y su longitud de onda eso entre los 400 y 7 00 nm. (Espectro, visible del espectro electromagnético), toda radiación cuya longitud de onda que eso fuera del rango mencionado no puede llamarse Luz de ahí que el uso de "luz" ultravioleta o infrarroja es inadecuado. Se debe nombrar como radiación ultravioleta (RUV) o infrarroja (RIR).
La radiación solar que llega al planeta tiene una amplia gama de longitudes de onda y en su conjunto se conoce como espectro electromagnético y la radiación lumínica está entre los 400 y 700 nm el resto de radiaciones forman parte de la porción no lumínica. Dentro de las radiaciones no lumínicas o no visibles, la radiación ultravioleta es la de mayor interés en fotodermatología. Tiene 3 bandas: el segmento A (UVA) entre los 320 a 400 nm; el segmento B (UVB) entre los 290 y 320 nm; y el segmento C (UVC) con longitudes de onda por debajo de 290 nm.
La radiación solar que llega al planeta tiene una amplia gama de longitudes de onda y en su conjunto se conoce como espectro electromagnético y la radiación lumínica está entre los 400 y 700 nm el resto de radiaciones forman parte de la porción no lumínica. Dentro de las radiaciones no lumínicas o no visibles, la radiación ultravioleta es la de mayor interés en fotodermatología. Tiene 3 bandas: el segmento A (UVA) entre los 320 a 400 nm; el segmento B (UVB) entre los 290 y 320 nm; y el segmento C (UVC) con longitudes de onda por debajo de 290 nm.
Otro aspecto importante a recordar es que cuando hablamos de Estados de Energía de la materia nos estamos refiriendo a los estados electrónicos de sus componentes (moléculas, átomos), caracterizados por su particular distribución en pares, contenido de energía y reactividad química singular. Normalmente las moléculas biológicas se encuentran en el menor estado de energía electrónica y reactividad química, estado que se conoce como Estado Electrónico Básico o Fundamental, llamándose a los estados de mayor energía causados por la absorción de un fotón, Estado Electrónico Excitado. Esta promoción de la molécula a estados de mayor energía se conoce como Transición y la desactivación de los estados electrónicos excitados que vuelven a su nivel de menor energía se conoce como Relajación. Los estados de excitación son llamados "Singlet" y "Triplet", ésta última de menor nivel de energía pero mas estable lo que le da mayor oportunidad de interactuar con otras moléculas del medio, permitiendo que las reacciones fotoquímicas secundarias ocurran. Hay que señalar que solo algunos tipos de moléculas químicas son capaces de ir a un estado triplet y estas son las moléculas que se saben que fluorescen. Como se aprecia, los estados moleculares electrónicamente excitados son inestables y si no ocurre cambio químico, se relajan. La relajación puede hacerse por un proceso radiante con emisión de un fotón; o a través de un proceso de no radiación como es la emisión de calor, de fluorescencia en el caso del decaimiento de energía del estado singlet o fosforescencia en caso del estado triplet, conversión Inter sistemas (del estado singlet al triplet) o por reacción química con las moléculas vecinas (Reacción Fotoquímica Secundaria). La Reacción Fotoquímica Primaria está dada por la absorción del fotón por el cromóforo.
Es importante recordar las leyes que rigen la interacción de radiación lumínica y materia:
- Ley de absorción fotoquímica o de Grotthus Draper o primera ley.- Solo las radiaciones que son absorbidas son efectivas para promover los cambios fotoquímicos.
- Ley de reciprocidad o de Burlen Roscoe o segunda ley.- Cuando el producto de la intensidad y el tiempo de exposición es consistente, el hecho fotoquímico es el mismo.
- Ley de absorción de Lambert Beer.- La fracción de luz incidente absorbida por una sustancia en solución es independiente de la intensidad de la luz inicial y aumenta proporcionalmente con el aumento en concentración de la sustancia.
- Ley del Quantum o de Planck.- La radiación es emitida no continuamente, sino en pequeñas unidades llamadas "quanta".
- Ley de equivalencia fotoquímica o de Starck Einstein.- Cuando un quantum de energía es absorbido por una molécula de una sustancia absorbente, se produce una activación de la molécula (reacción primaria).
Hay dos tipos básicos de reacciones fotoquímicas: Directas y por Fotosensibilización. Las Directas son aquellas en las que el cromóforo es afectado y sufre una modificación química para formar productos (pasaje de cis- a trans en el caso del ac. urocánico, por ejemplo) o también puede reaccionar con una molécula diferente como es el caso de la ciclo adición de los psoralenos bifuncionales con la timina del ADN. Las reacciones de Fotosensibilización, en las que el cromóforo solo juega el rol de captador y transportador de energía, más él no está afectado directamente por la reacción fotoquímica. En el extremo de la cadena él será restituido íntegramente y podrá de nuevo captar los fotones y reiniciar el proceso. Este tipo de cromóforo se llama "fotosensibilizador".
En el plano físico químico se distinguen dos tipos de reacción de fotosensibilización:
En el plano físico químico se distinguen dos tipos de reacción de fotosensibilización:
- Por transferencia de energía.- La energía del fotosensibilizador es transferida a un aceptador que pasa al estado de excitación. Un ejemplo es la reacción fotodinámica de tipo II.
- Por transferencia de carga o de un átomo de hidrógeno.- El fotosensibilizador excitado presenta propiedades de oxidorreducción y transfiere carga a un sustrato. Un ejemplo es la reacción fotodinámica de tipo I.
FOTOBIOLOGÍA CUTÁNEA
La Fotobiología Dermatológica es por tanto la parte de la Fotobiología que se ocupa del estudio de los cambios en la piel por efecto de la radiación no ionizante ambiental, especialmente de la radiación solar, y dentro del espectro solar, de la fracción ultravioleta y luz visible.
Es necesario conocer que la atmósfera filtra la radiación electromagnética solar en forma tal que sobre la superficie de la Tierra el espectro solar sólo abarca las longitudes de onda comprendidas entre las 290 y 3000 nm, es decir, la fracción UVB (290-320 nm) y UVA (320-400 nm), la luz visible (400-780nm) y una parte de la infrarroja (RIR).En la banda de RUV la UVB es 5 al 10% del total y el resto es UVA.
La calidad de la radiación directa que recibe la superficie del planeta varía según la altitud (la cantidad de UVB es 20% mayor a 1500 m de altitud que la reportada a nivel del mar), la latitud, la estación climática (en nuestra latitud es mayor en febrero, en Europa es en julio), la hora del día (entre las 11 y 15 horas es particularmente rico en nuestra latitud), la humedad y la polución atmosférica.
A esta radiación directa, cuando se considera al individuo, hay que sumar las radiaciones difuminadas por las partículas en suspensión de la atmósfera; así en atmósfera nubosa los altos cúmulos absorben la mayor parte de la radiación UV. Además hay que considerar la radiación reflejada por la nieve, 80%; por la arena, 20%; por el agua, 5%; por el césped, 3%. En total, la energía solar que llega a la superficie se reparte de la siguiente forma: 50% en RIR, 40% en luz y 10% en RUV siendo la porción RUV la más activa biológicamente.
Interacción de la piel con la radiación solar.
La piel interactúa con la radiación solar dando protección al ser vivo de la radiación biológicamente importante que son la RUV y la luz. Esta función la cumple gracias a su constitución física que le confiere determinadas propiedades ópticas y a las moléculas absorbentes de energía llamados cromóforos.
PROPIEDADES ÓPTICAS DE LA PIEL
Se rigen por cuatro procesos fundamentales:
La Fotobiología Dermatológica es por tanto la parte de la Fotobiología que se ocupa del estudio de los cambios en la piel por efecto de la radiación no ionizante ambiental, especialmente de la radiación solar, y dentro del espectro solar, de la fracción ultravioleta y luz visible.
Es necesario conocer que la atmósfera filtra la radiación electromagnética solar en forma tal que sobre la superficie de la Tierra el espectro solar sólo abarca las longitudes de onda comprendidas entre las 290 y 3000 nm, es decir, la fracción UVB (290-320 nm) y UVA (320-400 nm), la luz visible (400-780nm) y una parte de la infrarroja (RIR).En la banda de RUV la UVB es 5 al 10% del total y el resto es UVA.
La calidad de la radiación directa que recibe la superficie del planeta varía según la altitud (la cantidad de UVB es 20% mayor a 1500 m de altitud que la reportada a nivel del mar), la latitud, la estación climática (en nuestra latitud es mayor en febrero, en Europa es en julio), la hora del día (entre las 11 y 15 horas es particularmente rico en nuestra latitud), la humedad y la polución atmosférica.
A esta radiación directa, cuando se considera al individuo, hay que sumar las radiaciones difuminadas por las partículas en suspensión de la atmósfera; así en atmósfera nubosa los altos cúmulos absorben la mayor parte de la radiación UV. Además hay que considerar la radiación reflejada por la nieve, 80%; por la arena, 20%; por el agua, 5%; por el césped, 3%. En total, la energía solar que llega a la superficie se reparte de la siguiente forma: 50% en RIR, 40% en luz y 10% en RUV siendo la porción RUV la más activa biológicamente.
Interacción de la piel con la radiación solar.
La piel interactúa con la radiación solar dando protección al ser vivo de la radiación biológicamente importante que son la RUV y la luz. Esta función la cumple gracias a su constitución física que le confiere determinadas propiedades ópticas y a las moléculas absorbentes de energía llamados cromóforos.
PROPIEDADES ÓPTICAS DE LA PIEL
Se rigen por cuatro procesos fundamentales:
Los cromóforos de la piel son moléculas específicas que al absorber la RUV y la visible, aseguran los efectos biológicos de éstas sobre la piel. Estas moléculas tienen un característico espectro de absorción que depende de su estructura molecular. Los cromóforos son "energizados" por absorción de radiación y este hecho puede llevar a una reorganización molecular y/o a una acción con las moléculas vecinas (reacciones fotoquímicas). Por tanto, ninguna reacción fotoquímica tiene lugar en la piel si la longitud de onda a la que es expuesta la persona no es específicamente absorbida por los cromóforos de la piel.
La piel posee cromóforos endógenos normales o fisiológicos como son el ADN, las proteínas que contienen gran cantidad de aminoácidos aromáticos, los esteroles, los liposomas que dan lugar a reacciones cutáneas obligatorias, las porfirinas y el ácido urocánico en el estrato corneo. La melanina es un absorbente de la radiación lumínica y es el recurso fundamental de protección del daño producido. Al absorber al fotón es convertida a estados excitados, los cuales en su mayor parte los pierde en forma de calor, pero algunas de las moléculas activadas juegan rol en el daño celular y es así que en las quemaduras solares las células tienen mucha melanina. Ella produce daño celular. Otra función fisiológica de la melanina es atrapar y desactivar estados excitados. Además pueden estar los cromóforos anormales o fotosensibilizadores que pueden llevar a consecuencias no esperadas. Las porfirinas expuestas a fuente lumínica apropiada de UVA se activan y reacciona con el oxígeno. El oxigeno en estado singlet reacciona con el lípido de la membrana celular y forma un peróxido lípido que produce daño en la membrana celular. El antraceno actúa sobre organelas citoplasmáticas.
Los efectos biológicos de la exposición a la radiación solar son consecuencia de reacciones fotoquímicas producidas por la interacción de los fotones con los cromóforos normales de la piel. Clásicamente se dividen en función del tiempo que tarda la aparición de estos efectos.
EFECTOS PRECOCES O AGUDOS
La piel posee cromóforos endógenos normales o fisiológicos como son el ADN, las proteínas que contienen gran cantidad de aminoácidos aromáticos, los esteroles, los liposomas que dan lugar a reacciones cutáneas obligatorias, las porfirinas y el ácido urocánico en el estrato corneo. La melanina es un absorbente de la radiación lumínica y es el recurso fundamental de protección del daño producido. Al absorber al fotón es convertida a estados excitados, los cuales en su mayor parte los pierde en forma de calor, pero algunas de las moléculas activadas juegan rol en el daño celular y es así que en las quemaduras solares las células tienen mucha melanina. Ella produce daño celular. Otra función fisiológica de la melanina es atrapar y desactivar estados excitados. Además pueden estar los cromóforos anormales o fotosensibilizadores que pueden llevar a consecuencias no esperadas. Las porfirinas expuestas a fuente lumínica apropiada de UVA se activan y reacciona con el oxígeno. El oxigeno en estado singlet reacciona con el lípido de la membrana celular y forma un peróxido lípido que produce daño en la membrana celular. El antraceno actúa sobre organelas citoplasmáticas.
Los efectos biológicos de la exposición a la radiación solar son consecuencia de reacciones fotoquímicas producidas por la interacción de los fotones con los cromóforos normales de la piel. Clásicamente se dividen en función del tiempo que tarda la aparición de estos efectos.
EFECTOS PRECOCES O AGUDOS
- Acción calórica debido a la fracción RIR que provoca vasodilatación cutánea responsable del eritema con elevación de la temperatura cutánea; por mecanismo reflejo la secreción sudoral asegura la regulación térmica. Una sobre exposición puede conducir al golpe de calor por saturación de las posibilidades de termorregulación, especialmente en los infantes.
- Pigmentación inmediata, directa, primaria o fenómeno de Meirowski, producido por la UVA aparece pocos minutos después de la exposición y dura pocas horas. La reacción fotoquímica parece ser una foto oxidación de la melanina.
- Acción antirraquítica de los UVB que inducen la transformación en la epidermis del 7-dehidrocalciferol en colecalciferol, el que a nivel del hígado y riñón será hidrolizado para formar el 1,25 dihidrocolecalciferol.
EFECTOS RETARDADOS
- El eritema actínico, de manifestación clínica tan conocida es producida por UVA. Su mecanismo de acción planteado está entre una acción de la RUV sobre los cromóforos dérmicos (lisosomas endoteliales, membrana de las células endoteliales con liberación de prostaglandinas y membranas de mastocitos peri vasculares con liberación de serotonina e histamina) y una acción indirecta por liberación de mediadores epidérmicos que se difunden hacia la dermis (las prostaglandinas parecen ser mediadores responsables de la vasodilatación).
- Pigmentación retardada o tardía, muy conocida, que se inicia dos días después de la exposición alcanzando su máximo a las 3 semanas y disminuye progresivamente. Su mecanismo de acción es por neosíntesis melánica.
- Hiperplasia epidérmica consecuencia de la interacción de los fotones con el ADN epidérmico. Después de la inhibición inicial sigue un aumento en las mitosis de los queratinocitos a partir de las 48 horas, conduciendo al engrosamiento de la capa cornea. Esta es interpretada como una reacción de fotoprotección.
EFECTOS CRÓNICOS
La dosis de irradiación lumínica recibida por la piel es acumulativa. La aparición de los efectos crónicos dependen tanto de la forma en que las dosis son recibidas como de la fotoprotección natural del individuo.
La dosis de irradiación lumínica recibida por la piel es acumulativa. La aparición de los efectos crónicos dependen tanto de la forma en que las dosis son recibidas como de la fotoprotección natural del individuo.
- Envejecimiento cutáneo precoz o fotoenvejecimiento. Esto es particularmente visible en zonas expuestas (nuca, dorso de manos, frente, pómulos). Actualmente se da la eficacia espectral máxima al UVB y luego al UVA. La RIR parece potenciar el efecto de las RUV al menos en la elastosis.
- Fotocarcinogénesis. El rol de la radiación no iónica sobre la carcinogénesis cutánea no melanoma, esta claramente establecida con argumentos clínicos y epidemiológicos como son la localización en áreas cutáneas más expuestas a la luz, mayor frecuencia de presentación en los que realizan trabajos en áreas abiertas, en sujetos con fotosensibilidad especial (xerodermia pigmentosa, albinismo). La frecuencia se duplica cuando uno se desplaza hacia el Ecuador y se multiplica por 10 en las personas de fototipo I y II que viven en Australia, comparada con personas del mismo fototipo de piel que viven en Gran Bretaña. Además experimentalmente se ha establecido plenamente en el ratón. La relación es dosis dependiente a la dosis acumulativa de UV en el curso de la vida. Las exposiciones precoces jugarían un papel agravante. La RUVB tiene acción casi directa sobre carcinogenésis, la UVA tiene papel inductor La luz y la RIR tendrían un efecto promotor. Su mecanismo íntimo no está bien establecido. En lo que respecta al melanoma maligno cutáneo (MMC) el rol oncogénico del sol no esta bien establecido. Hay ciertos hechos que argumentan en contra, tales como la ausencia de correlación entre la dosis requerida y la aparición de melanoma, la localización frecuente en zonas cubiertas, su aparición a una edad más precoz, observarse más en áreas urbanas y la falta de elastosis solar asociada a MMC. Pero sí podemos decir que el MMC es inducido por la exposición solar en razón de los siguientes hechos: 1) antecedentes de exposiciones intensas y traumáticas (por ej: las observadas en las vacaciones de verano) y el de exposiciones precoces en la vida; 2) La mayor frecuencia de MMC en las pieles de tipos I y II; 3) el melanoma léntigo maligno está localizado casi exclusivamente en zonas expuestas.
Foto inmunología o efectos inmunológicos del Sol.
Kripke, en trabajos experimentales realizados en ratón ha demostrado que algunas respuestas inmunes pueden ser inhibidas por la exposición a la RUV abriendo una senda de investigación aún en explotación. Se ha demostrado que la RUV particularmente la UVB aplicada en forma repetida en ratones inhiben el rechazo de tumores fotoinducidos que les han sido transplantados. Estos tumores fuertemente antigénicos son rechazados cuando se transplantan a ratones no radiados. Esta tolerancia es especifica al tumor y puede ser transferida por inyección de células linfoides provenientes de ratón singénico irradiado. Así mismo se observa que la dosis simple de UVB aplicada a ratones inhibe el desarrollo de hipersensibilidad local por contacto a una sustancia química simple. A dosis mayores; puede producir supresión sistémica de la hipersensibilidad por contacto.
Tanto la tolerancia provocada para tumores fotoinducidos, como la inhibición de la reacción de hipersensibilidad retardada, son debidas a la generación de linfocitos T supresores antígeno específicos.
Los mecanismos de estas dos modificaciones inmunológicas no están claras. Es posible que la irradiación altere la función de los macrófagos involucrados en el procesamiento y presentación de antígenos, especialmente en las células de Langerhans integrantes de la epidermis y que constituye el 4% de su población celular. Hay indicios que la UVC mata a la células de Langerhans; que las UVB son mejores supresores y que las UVA inducen. Esta interacción de la radiación no ionizante con el sistema inmune esta involucrada en la patogenia de algunas enfermedades como la fotoalergia y la urticaria solar Es conocida la exacerbación de enfermedades como el lupus eritematoso, el pénfigo y el penfigoide por radiación solar, así como el efecto benéfico del PUVA y UVB en el tratamiento de la micosis fungoides, liquen plano, vitiligo y otras enfermedades.